Hace unos cincuenta años, dos años después del descubrimiento de la estructura del ADN, todos suponían que la estructura de un gen era una sola cadena de pares de bases, desde donde la cual se transfiere la información para la síntesis de una proteína. Esta suposición era en gran parte correcta para el reino de los organismos procarióticos, pero años más tarde hubo indicios de que estos conceptos simples para los genes podrían no explicar algunas observaciones en los organismos eucarióticos.
En los 1970s, otra observación desconcertante fue la detección de ARN inusualmente largo en el núcleo de las células de vertebrados en comparación con el ARNm más corto que emergía en el citoplasma. Sorprendentemente, este ARN nuclear tenía una estructura de caperuza en su extremo 5′ y un tramo de poliadenosina [poli(A)] en su extremo 3′, al igual que el ARNm citoplasmático más corto. La resolución de estas paradojas ocurrió con el descubrimiento de genes divididos y empalme de ARN.
Una simple comparación de la secuencia de un ARNm y su ADN nuclear correspondiente reveló secuencias que se eliminaron mediante empalme durante el procesamiento del precursor más largo. Esto explicó de inmediato cómo el ARN nuclear largo y el ARNm citoplasmático más corto podían tener los mismos extremos, un casquete y una cola de poli(A), mientras que las diferencias en longitud se debían a la eliminación de secuencias de intrones del medio.
Pronto se demostró que casi todos los genes en los organismos multicelulares contenían intrones, con un promedio de diez por gen en los vertebrados. Además, la presencia de estos intrones representó una responsabilidad genética para los genes;
Aproximadamente el 25% de todas las mutaciones en los genes de la globina que causan la β-talasemia en humanos se debieron a defectos en el corte y empalme. La comparación de las secuencias de intrones y exones (los nombres sugeridos por Wally Gilbert) reveló la presencia de secuencias de consenso en sus límites y la mutación de estas secuencias inactivaron el empalme.
Introducción al empalme de genes
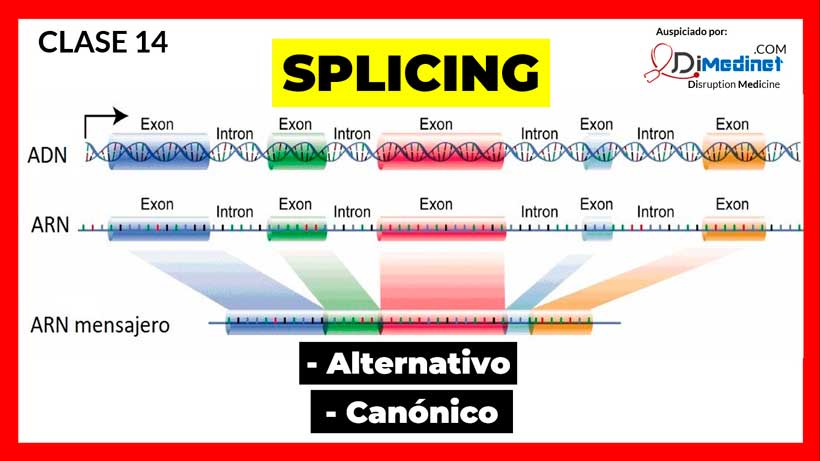
El empalme de genes es una modificación postranscripcional en la que un solo gen puede codificar múltiples proteínas. El empalme de genes se realiza en eucariotas, antes de la traducción del ARNm, mediante la inclusión o exclusión diferencial de regiones de pre-ARNm. El empalme de genes es una fuente importante de diversidad de proteínas. Durante un evento típico de empalme de genes, el pre-ARNm transcrito de un gen puede conducir a diferentes moléculas de ARNm maduras que generan múltiples proteínas funcionales.
Por lo tanto, el empalme de genes permite que un solo gen aumente su capacidad de codificación, lo que permite la síntesis de isoformas de proteínas que son estructural y funcionalmente distintas. El empalme de genes se observa en una alta proporción de genes. En las células humanas, se sabe que alrededor del 40-60% de los genes exhiben empalmes alternativos.
Mecanismo de empalme de genes
Hay varios tipos de eventos comunes de empalme de genes. Estos son los eventos que pueden ocurrir simultáneamente en los genes después de que se forma el ARNm a partir del paso de transcripción del dogma central de la biología molecular.
Omisión de exón: este es el mecanismo de empalme de genes conocido más común en el que los exones se incluyen o excluyen de la transcripción del gen final que conduce a variantes de ARNm extendidas o acortadas. Los exones son las regiones codificantes de un gen y son responsables de producir proteínas que se utilizan en varios tipos de células para una serie de funciones.
Retención de intrones: un evento en el que un intrón se retiene en la transcripción final. En humanos, se ha informado que del 2 al 5 % de los genes retienen intrones. El mecanismo de empalme de genes retiene las porciones no codificantes (basura) del gen y conduce a una degradación de la estructura y la funcionalidad de la proteína.
Sitio de empalme 3′ y sitio de empalme 5′ alternativos: el empalme alternativo incluye la unión de diferentes sitios de empalme 5′ y 3′. En este tipo de empalme de genes, dos o más sitios de empalme 5′ alternativos compiten por unirse a dos o más sitios de empalme 3′ alternativos.
Métodos de detección de variantes de empalme
Actualmente, los métodos más eficientes para la detección a gran escala de variantes de empalme incluyen métodos de predicción computacional y análisis de micromatrices. La detección de variantes de empalme basada en micromatrices es el método más popular actualmente en uso. La naturaleza altamente paralela y sensible de los microarreglos los hace ideales para monitorear la expresión génica en un nivel de genoma completo específico de tejido.
Los métodos basados en micromatrices para detectar variantes de empalme proporcionan una plataforma robusta para el descubrimiento de alto rendimiento de empalmes de genes alternativos. Se detectaron varios transcritos de genes novedosos usando métodos basados en micromatrices que no fueron detectados por EST usando métodos computacionales.
Otro método comúnmente utilizado para descubrir nuevas isoformas de genes es la RT-PCR seguida de la secuenciación. Este es un enfoque poderoso y puede usarse de manera efectiva para analizar una pequeña cantidad de genes. Sin embargo, solo proporciona una visión limitada de la estructura del gen, requiere mucha mano de obra y no se escala fácilmente a miles de genes o cientos de tejidos.
El pasado y el futuro de los intrones
La existencia de intrones y empalmes diferenciales ayuda a explicar cómo se crean nuevos genes durante la evolución. El empalme hace que los genes sean más “modulares”, lo que permite que se creen nuevas combinaciones de exones durante la evolución. Además, se pueden insertar nuevos exones en intrones antiguos, creando nuevas proteínas sin interrumpir la función del gen antiguo.
Nuestro conocimiento del empalme de ARN es bastante nuevo. No obstante, debido a que casi todos los eucariotas tienen intrones y comparten mecanismos de empalme de ARN, el empalme en sí debe ser bastante antiguo. Los defensores de la teoría del “intrón temprano” sugieren que todos los organismos (incluidos los procariotas) alguna vez tuvieron intrones en su genoma, pero posteriormente perdieron estos elementos, mientras que los partidarios del “intrón tardío” creen que la restricción de los intrones a los eucariotas sugiere una teoría más reciente. introducción (Roy y Gilbert, 2006).
No existe un patrón aparente en el que los eucariotas tengan intrones, y eso dificulta que los investigadores hagan predicciones sobre cómo se ganaron o perdieron intrones a lo largo de la evolución. Sin embargo, lo que está claro es que los intrones y el empalme han desempeñado claramente un papel importante en la evolución.
En este BioCast que te preparamos, te explico sobre cuales son los elementos que aparecen en la maduración del ARN mensajero antes del proceso de splicing.
Resumen minuto a minuto lo que aprenderás en este video:
00:00 Intro
00:20 Qué es el SPLICING
00:22 Ribonucleoproteínas que participan
01:10 Splicing canónico
02:12 El spliceosome
02: 26 Splicing alternativo
04: 35 Ejemplos de splicing alternativo
05:03 Splicing en el gen DSCAM de la drosophila
07:28 Splicing en el gen de la apolipoproteína b
Como siempre acepto comentarios y sugerencias.
Edwin W. editor en DiMedinet y Bits de Ciencia.
 (Ninguna valoración todavía)
(Ninguna valoración todavía)![]() Cargando...
Cargando...